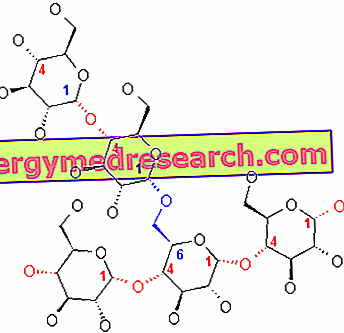
Το γλυκογόνο είναι ένα μακρομόριο (μοριακής μάζας περίπου 400 εκατομμυρίων daltons) α-γλυκόζης στο οποίο υπάρχουν κυρίως α-1, 4 γλυκοσιδικούς δεσμούς και κλάδους σε αναλογία 1:10, λόγω α-1, 6 γλυκοσιδικών δεσμών.
Το γλυκογόνο συνιστά εφεδρικό υλικό και αποικοδομείται συνεχώς και ανασυστάται. σε όλη τη μάζα των κυττάρων του σώματος, έχουμε περίπου 100 g γλυκογόνου: το μεγαλύτερο μέρος του είναι στο ήπαρ όπου είναι κινητό και μπορεί ως εκ τούτου να χρησιμοποιηθεί ως αποθεματικό για τα άλλα όργανα (το γλυκογόνο στους μυς δεν είναι κινητό).
Τα ένζυμα που καταλύουν την αποικοδόμηση και τη σύνθεση του γλυκογόνου είναι όλα στο κυτταρόπλασμα και επομένως χρειάζεται ένα σύστημα ρύθμισης που καθιστά μια διαδρομή αδρανή όταν η άλλη είναι ενεργή: εάν είναι διαθέσιμη η γλυκόζη, η τελευταία μετατρέπεται σε γλυκογόνο (αναβολισμός) που είναι ένα απόθεμα, αντίθετα, εάν απαιτείται γλυκόζη, τότε το γλυκογόνο υποβαθμίζεται (καταβολισμός).
Το ένζυμο που εμπλέκεται κυρίως στην αποικοδόμηση του γλυκογόνου είναι φωσφορυλάση γλυκογόνου . αυτό το ένζυμο είναι ικανό να διασπά έναν α-1, 4 γλυκοσιδικό δεσμό χρησιμοποιώντας, ως λυτικό παράγοντα, ένα ανόργανο ορθοφωσφορικό: η διάσπαση λαμβάνει χώρα μέσω φωσφορολυτικού και λαμβάνεται 1-φωσφορική γλυκόζη.
Σε πέντε ή έξι μονάδες από ένα σημείο διακλάδωσης, το ένζυμο φωσφορυλάση γλυκογόνου δεν είναι πλέον ικανό να δράσει, επομένως αποσπάται από το γλυκογόνο και αντικαθίσταται από ένα ένζυμο αποκοπής που είναι μια τρανσφεράση : στην καταλυτική θέση αυτού του ενζύμου υπάρχει «ιστιδίνη που επιτρέπει τη μεταφορά τριών σακχαριτικών μονάδων στην πλησιέστερη γλυκοσιδική αλυσίδα (η ιστιδίνη επιτίθεται στον πρώτο άνθρακα ενός μορίου γλυκόζης). Το ένζυμο που μόλις αναφέρθηκε είναι γλυκοσυλοτρανσφεράση . στο τέλος της δράσης αυτού του ενζύμου, μόνο μία μονάδα γλυκόζης παραμένει στην πλευρική αλυσίδα με τον πρώτο άνθρακα δεσμευμένο στον έκτο άνθρακα μιας γλυκόζης στην κύρια αλυσίδα. Η τελευταία μονάδα γλυκόζης της πλευρικής αλυσίδας απελευθερώνεται από τη δράση του ενζύμου α-1, 6 γλυκοσιδάσης (αυτό το ένζυμο είναι το δεύτερο μέρος του ενζύμου αποκοπής). δεδομένου ότι οι κλάδοι στο γλυκογόνο είναι σε αναλογία 1:10, από την πλήρη αποικοδόμηση του μακρομορίου λαμβάνουμε περίπου 90% 1-φωσφορική γλυκόζη και περίπου 10% γλυκόζη.
Η δράση των προαναφερθέντων ενζύμων επιτρέπει την απομάκρυνση μιας πλευρικής αλυσίδας από το μόριο γλυκογόνου. η δραστικότητα αυτών των ενζύμων μπορεί να επαναληφθεί μέχρι να υπάρξει πλήρης αποικοδόμηση της αλυσίδας.
Ας εξετάσουμε ένα ηπατοκύτταρο. γλυκόζη (αφομοιωμένη μέσω της διατροφής), όταν εισέρχεται στο κύτταρο μετατρέπεται σε 6-φωσφορική γλυκόζη και έτσι ενεργοποιείται. Η 6-φωσφορική γλυκόζη, με τη δράση της φωσφογλυκομουτάσης, μετατρέπεται σε 1-φωσφορική γλυκόζη: η τελευταία είναι ένας μη άμεσος πρόδρομος της βιοσύνθεσης. σε βιοσύνθεση, χρησιμοποιείται μια ενεργοποιημένη μορφή σακχάρων, η οποία αντιπροσωπεύεται από ζάχαρη συνδεδεμένη με ένα διφωσφορικό: συνήθως ουριδινυλοδιφωσφορική (UDP). Το 1-φωσφορικό γλυκόζη στη συνέχεια μετατρέπεται σε UDP-γλυκόζη. αυτός ο μεταβολίτης είναι υπό τη δράση της συνθάσης γλυκογόνου η οποία είναι ικανή να δεσμεύει την UDP-γλυκόζη σε ένα μη αναγωγικό άκρο του αναπτυσσόμενου γλυκογόνου: επιμηκυμένο γλυκογόνο λαμβάνεται από μία γλυκοσιδική μονάδα και UDP. Το UDP μετατρέπεται από το ένζυμο nucleosidedifosfokinase σε UTP που επιστρέφει στην κυκλοφορία.
Η αποικοδόμηση του γλυκογόνου συμβαίνει με τη δράση της φωσφορυλάσης γλυκογόνου η οποία απελευθερώνει ένα μόριο γλυκόζης και μετασχηματίζει το σε 1-φωσφορική γλυκόζη. Στη συνέχεια, η φωσφογλυκομουτάση μετατρέπει την 1-φωσφορική γλυκόζη σε 6-φωσφορική γλυκόζη.
Το γλυκογόνο συντίθεται πάνω από όλα στο ήπαρ και στους μύες: στο σώμα υπάρχουν 1-1, 2 γραμμάρια γλυκογόνου που κατανέμονται σε ολόκληρη τη μυϊκή μάζα.
Το γλυκογόνο ενός μυοκυττάρου αντιπροσωπεύει ένα απόθεμα ενέργειας μόνο για αυτό το κύτταρο, ενώ το γλυκογόνο που περιέχεται στο ήπαρ είναι επίσης ένα απόθεμα για τους άλλους ιστούς, δηλαδή μπορεί να αποσταλεί, ως γλυκόζη, σε άλλα κύτταρα.
Η 6-φωσφορική γλυκόζη που λαμβάνεται στους μυς από την αποικοδόμηση του γλυκογόνου στη συνέχεια αποστέλλεται, στην περίπτωση ενεργειακών απαιτήσεων, στη γλυκόλυση. στο ήπαρ, η 6-φωσφορική γλυκόζη μετατρέπεται σε γλυκόζη με τη δράση φωσφατάσης 6-φωσφορικής γλυκόζης (ένα ένζυμο χαρακτηριστικό των ηπατοκυττάρων) και μεταφέρεται στο κυκλοφοριακό ρεύμα.
Τόσο η συνθετάση γλυκογόνου όσο και η γλυκογόνο φωσφορυλάση δουλεύουν στις μη αναγωγικές μονάδες γλυκογόνου, οπότε πρέπει να υπάρχει ένα ορμονικό σήμα που να διευθύνει την ενεργοποίηση μίας οδού και την παρεμπόδιση του άλλου (ή αντίστροφα).
Στο εργαστήριο ήταν δυνατό να επιμηκυνθεί η αλυσίδα του γλυκογόνου, εκμεταλλευόμενη φωσφορυλάση γλυκογόνου και χρησιμοποιώντας 1-φωσφορική γλυκόζη σε πολύ υψηλή συγκέντρωση.
Στα κύτταρα, φωσφορυλάση γλυκογόνου καταλύει μόνο την αντίδραση αποικοδόμησης επειδή οι συγκεντρώσεις των μεταβολιτών είναι τέτοιες ώστε να μετατοπίζουν την ισορροπία της ακόλουθης αντίδρασης, προς τα δεξιά (δηλ. Προς την αποικοδόμηση του γλυκογόνου):

Ας δούμε τον μηχανισμό δράσης της φωσφορυλάσης γλυκογόνου: το ακεταλικό οξυγόνο (το οποίο δρα ως γέφυρα μεταξύ των μονάδων γλυκόζης) συνδέεται με το φωσφορυλικό υδρογόνο: ένα ενδιάμεσο της αντίδρασης σχηματίζεται από ένα carbocatione (σε γλυκόζη που είναι άκρο) στην οποία δεσμεύεται πολύ γρήγορα το φωσφορυλικό (Pi).
Η φωσφορυλάση γλυκογόνου απαιτεί έναν συμπαράγοντα ο οποίος είναι φωσφορική πυριδοξάλη (αυτό το μόριο είναι επίσης συμπαράγοντας για τις τρανσαμινάσες): έχει πρωτονιωμένο φωσφορυλ μόνο εν μέρει (το φωσφορικό πυριδοξάλη περιβάλλεται από ένα υδρόφοβο περιβάλλον που δικαιολογεί την παρουσία πρωτονίων που δεσμεύονται σε αυτό) . Το φωσφορυλ (Pi) είναι ικανό να αποδώσει ένα πρωτόνιο στο γλυκογόνο επειδή ένα τέτοιο φωσφορυλ ανακτά τότε το πρωτόνιο από το μερικώς πρωτονιωμένο φωσφορυλ του φωσφορικού πυριδοξάλης. Η πιθανότητα ότι, στο φυσιολογικό ρΗ, το φωσφόρο χάνει το πρωτόνιο και παραμένει εντελώς αποπρωτονιωμένο είναι πολύ χαμηλό.
Τώρα ας δούμε πώς λειτουργεί το phosphoglucomutase. Αυτό το ένζυμο έχει φωσφορυλιωμένο υπόλειμμα σερίνης στην καταλυτική θέση. η σερίνη αποδίδει φωσφορυλ σε 1-φωσφορική γλυκόζη (στη θέση έξι): 1, 6-διφωσφορικό γλυκόζη σχηματίζεται για μικρό χρονικό διάστημα, στη συνέχεια η σερίνη αναφωσφορυλιώνεται με λήψη φωσφορυλίου στη θέση ένα. Η μετάλλαξη Phosphoglucus μπορεί να λειτουργήσει και στις δύο κατευθύνσεις, δηλ. Να μετατρέψει 1-φωσφορική γλυκόζη σε 6-φωσφορική γλυκόζη ή αντίστροφα. εάν παράγεται 6-φωσφορική γλυκόζη, μπορεί να αποσταλεί απευθείας στη γλυκόλυση, στους μύες ή να μετατραπεί σε γλυκόζη στο ήπαρ.
Το ένζυμο ουριδίλ φωσφογλυκό τρανσφεράση (ή UDP γλυκόζης πυροφωσφορυλάση) καταλύει την αντίδραση μεταφοράς της 1-φωσφορικής γλυκόζης σε UTP με επίθεση φωσφορυλίου α.
Το ένζυμο που μόλις περιγράψαμε είναι πυροφωσφορυλάση: αυτό το όνομα οφείλεται στο γεγονός ότι η αντίθετη αντίδραση με αυτή που μόλις περιγράφηκε είναι πυροφωσφορυλίωση.
Η UDP γλυκόζη, που λαμβάνεται όπως περιγράφεται, είναι ικανή να επιμηκύνει την αλυσίδα γλυκογόνου μιας μονάδας μονοσακχαρίτη.
Είναι δυνατόν να εξελιχθεί η αντίδραση προς τον σχηματισμό γλυκόζης UDP εξαλείφοντας ένα προϊόν το οποίο είναι πυροφωσφορικό. το ένζυμο πυροφωσφατάση μετατρέπει το πυροφωσφορικό σε δύο ορθοφωσφορικά μόρια (υδρόλυση ενός ανυδρίτη) και με αυτό τον τρόπο διατηρεί τη συγκέντρωση του πυροφωσφορικού τόσο χαμηλή ώστε η διαδικασία σχηματισμού γλυκόζης UDP προτιμάται θερμοδυναμικά.
Όπως αναφέρθηκε, η UDP γλυκόζη, χάρη στη δράση της συνθάσης γλυκογόνου, είναι ικανή να επιμηκύνει την αλυσίδα του γλυκογόνου.
Οι διακλαδώσεις (σε αναλογία 1:10) οφείλονται στο γεγονός ότι, όταν μια αλυσίδα γλυκογόνου αποτελείται από 20-25 μονάδες, παρεμβαίνει ένα ένζυμο διακλάδωσης (που έχει μια ιστιδίνη στην καταλυτική του θέση) ικανό να μεταφέρει μια σειρά 7 -8 γλυκοσιδικές μονάδες περαιτέρω προς τα κάτω των 5-6 μονάδων: δημιουργείται έτσι μια νέα διακλάδωση.
Για λόγους νευρικής προέλευσης ή εάν απαιτείται ενέργεια λόγω σωματικής προσπάθειας, η αδρεναλίνη εκκρίνεται από τα επινεφρίδια.
Τα κύτταρα στόχοι της αδρεναλίνης (και της νορεπινεφρίνης) είναι αυτά του ήπατος, των μυών και του λιπώδους ιστού (στην τελευταία υπάρχει η αποικοδόμηση των τριγλυκεριδίων και η κυκλοφορία των λιπαρών οξέων: στα μιτοχόνδρια παράγεται επομένως η γλυκόζη 6 το φωσφορικό άλας της γλυκόζης μετασχηματίζεται σε γλυκόζη με τη δράση του ενζύμου φωσφατάση 6-φωσφορικής γλυκόζης και εξάγεται στους ιστούς).
Ας δούμε τώρα τον τρόπο δράσης της αδρεναλίνης. Η αδρεναλίνη συνδέεται με έναν υποδοχέα τοποθετημένο στην κυτταρική μεμβράνη (των μυοκυττάρων και των ηπατοκυττάρων) και αυτό καθορίζει τη μετάφραση του σήματος από έξω προς το εσωτερικό του κυττάρου. Η πρωτεϊνική κινάση ενεργοποιείται η οποία δρα ταυτόχρονα στα συστήματα που ρυθμίζουν τη σύνθεση και αποικοδόμηση του γλυκογόνου:
Η συνθετάση γλυκογόνου υπάρχει σε δύο μορφές: μία αποφωσφορυλιωμένη μορφή (ενεργή) και μία φωσφορυλιωμένη μορφή (ανενεργή). η πρωτεϊνική κινάση φωσφορυλιώνει τη συνθάση του γλυκογόνου και αποκλείει τη δράση της.
Η φωσφορυλάση γλυκογόνου μπορεί να υφίσταται σε δύο μορφές: μία ενεργή μορφή στην οποία υπάρχει μία φωσφορυλιωμένη σερίνη και μία αδρανής μορφή στην οποία η σερίνη είναι αποφωσφορυλιωμένη. Η φωσφορυλάση γλυκογόνου μπορεί να ενεργοποιηθεί από το ένζυμο γλυκογόνο φωσφορυλάση κινάση . Η κινάση της φωσφορυλάσης του γλυκογόνου είναι δραστική εάν φωσφορυλιωθεί και αδρανής εάν είναι αποφωσφορυλιωμένη. η πρωτεϊνική κινάση έχει ως υπόστρωμα την κινάση γλυκογόνου φωσφορυλάσης, δηλ. είναι ικανή να φωσφορυλιώνει (και επομένως να ενεργοποιεί) την τελευταία, η οποία, με τη σειρά της, ενεργοποιεί γλυκογόνο φωσφορυλάση.
Μόλις τελειώσει το σήμα αδρεναλίνης, το αποτέλεσμα που έχει στο κύτταρο πρέπει επίσης να τερματιστεί: τα ένζυμα φωσφατάσης παρεμβαίνουν, στη συνέχεια, στα είδη πρωτεϊνών.